by Guy Thompson, No.24 Summer 2022
An unfortunate result of our improved understanding of the causes of disease and the spread of disease is that we have been conditioned to regard all ‘germs’ – bacteria, viruses and fungi – as ‘bad’ and to believe that we need to keep ourselves (and children especially) in clean, sanitised environments away from these germs. The “Hygiene Hypothesis” was first put forward in the late 1980s by David Strachan, a professor of epidemiology, who had identified that children from larger households showed lower susceptibility to certain diseases and conditions, and suggested that the reason for this was that children with more siblings were exposed to more germs. He also noted that a reducing number of infections in western countries correlated with an increase in the number of cases of auto-immune and allergic disease and concluded that an individual’s susceptibility to disease went up if they were raised in more sanitised environments. He was the first to suggest that exposure to germs and bacteria is not a bad thing as it helps develop and sustain the immune system.
The thinking behind the Hygiene Hypothesis is fairly straightforward. When born, babies emerge from a sterile environment in the womb and normally have had no – or at least very limited – exposure to pathogens. As a result, their innate immune system is undeveloped and they are highly susceptible to infection; only after being exposed to different bacteria and viruses does the immune system develop and strengthen. This has been thought to explain why, for example, having a pet animal in the home results in better health and fewer allergies in later life. However, the Hygiene Hypothesis has been unable to provide a complete answer. Allergic asthma, for example, is on the increase in many “unhygienic” cities in America, and migrants in some large European cities show a noticeably lower incidence of allergic disease than the locals; under the Hygiene Hypothesis the reverse would be expected. With these inconsistencies in mind, the “Old Friends” hypothesis was put forward in 2003 by Prof. Graham Rook who argued that some microbes that have been around throughout human history and that have therefore been constantly present during the evolution of the human immune system are so entwined with those systems that they have come to form an essential part of it. Early exposure to a diverse range of “friendly” microbes – not infectious pathogens – is needed to train the human immune system to react appropriately. In an increasingly germ-free world, this rela- tionship is broken [1].
More than 90% of human evolution is believed to have taken place as isolated hunter-gatherers and, later, in early farming communities, with high levels of exposure to soil, funguses and rotting vegetation. Given this, the evolution of the immune system was conditioned by the presence of certain microbes. Indeed, some microbes have co-evolved alongside the human immune system to the extent that it is impossible for one to function properly without the other – hence the term “Old Friends”. Unlike the hygiene hypothesis, where the presence of bacteria or germs is considered good for stimulating immune response, the Old Friends approach indicates quite clearly that certain types of bacteria are actually a functioning part of the immune system and truly necessary for its proper operation. So, under the Old Friends Hypothesis, the incidence of childhood disease may be explained by a lack of timely exposure to the right strains of bacteria; in later life, allergies, weight gain, chronic diseases, etc., are similarly a function of an earlier or ongoing deficiency. With Old Friends, the immune system ceases to be regarded as stand-alone – challenged and thereby primed to provide for our protection – but rather as founded upon that early input of microbiota, an input which today is so often either missing as we reduce our exposure to the natural environment, or radically changed from the types of microbes we used to encounter in the period the immune system evolved.
Although the two hypotheses are not mutually exclusive, the growing improvement in our understanding means that, quoting from Prof. Rook, “we now know how the adaptive immune system and the complex microbiota co-evolved, so that the immune system can “farm” the microbiota while at the same time exclude pathogens. The innate immune system was not sufficient for this purpose”. He continues that “without appropriate microbial inputs the regulation of the immune system is faulty, and the risk of chronic inflammatory disorders increases. These increases are already striking in high income settings, and are increasing globally as prosperity increases. This happens because there are lifestyle changes … that diminish the microbial inputs, and distort the microbiota/microbiome and lead to faulty immunoregulation.”
Our damaging obsession with the elimination of beneficial agents extends beyond human health into agriculture, where “health” is often defined as an absence of bacteria, viruses and parasites. As with humans, this compromises not just the very positive role of infectious microbes and parasites in maintaining health, but their absolute necessity for the early establishment of functioning immune protection. This applies particularly to insects where it is known that immune systems are generally simpler and thus, pro rata, more reliant on the extra contribution of Old Friends. When it comes to honey bees, work by Isobel Grindrod and Stephen Martin suggests that the disease elimination approaches common in modern bee farming may well create a substantial barrier to the development of a balanced relationship between pathogens and their hosts [2]. Untoward outcomes from insecticide use are well studied, but herbicides and fungicides cause just as great a problem for many insects as they directly attack the proper functioning of the gut micro-biota and destroy the very foundations of any evolved immune response. So for insects, with an equal or, proportionately, even greater reliance upon the availability of Old Friends, it may be that the functioning of their immune systems is a lot more similar to that of mammals than had been thought and a lot more exposed as the environment diminishes [3]. And as with humans where, for example, a Western diet lacking in plant fibre and other diverse foods appears to disrupt the healthy microbiota in our guts, the same is true of managed honey bees for which artificial feeding, colony level chemical treatments and regular fungicide exposure via foraging has become the norm.
Individual honey bees spend a significant portion of their lives as larvae and then pupae. If the Old Friends Hypothesis applies, it is reasonable to assume that, rather as with the umbilical cord in mammals, the early exposure to microbes necessary for development of the immune system occurs at these stages [4]. This might be through the supply of food and water – both of which originate in and reflect the local landscape – or by simple exposure to the mediated nest atmosphere. Even after emergence as a worker, there is normally no significant direct contact with the outside world for several weeks as the young bee works through the ranks within the nest. In the same way that children continue amassing microbiota in every contact with family members, while playing outside in dirt, getting licked by dogs, or sharing toys with friends, young honey bees’ developing immune systems take cues from all the encounters within the hive. By the time they are promoted to outdoor activities the immune system is developed and the crucial exposure to supportive microbes has taken place entirely within that controlled environment. There is therefore a substantial period where low levels of exposure to pathogens brought back to the nest by foraging bees interact with the individual immune systems and stimulate later protection against attack by higher virulence pathogens both in the nest and out in the field; this provides a real and lasting health benefit across the entire colony. Equally, a low but continuous exposure to parasites benefits the overall health of the colony and, by extension, the wider local community; it is therefore possible that the health and resilience of honey bee local populations could be as dependant on a healthy parasite community as it is on diverse forage, say, or a biodiverse fungal landscape. It must also be recognised that interrupting these interactions, as happens with almost any level of honey bee management, causes significant disruption to the establishment and maintenance of immunoregulation in the individual bee and the colony.

With honey bees we are not talking about a single layer of disease protection; the individual bees have their own immune systems but the colony superorganism adds a second layer of complexity [5]. The colony lives within a very substantial outer “skin” – usually in a living tree – that facilitates efficient environmental regulation, with temperature and humidity being very important for the development of brood within the nest. Perhaps more importantly, the outer wall allows the controlled maintenance of an antimicrobial atmosphere within the nest as part of an overall strategy to both control disease and regulate symbiotic interactions. This envelope is achieved with propolis; resins painstakingly collected, mixed with saliva and wax, and deposited all around the walls of the cavity. The energetic cost of doing this is very significant and, it can therefore be assumed, very important to the functioning of this quasi immune system. Interestingly, the flavonoid Galangin – a strong scavenger antioxidant found in manuka honey and a potential alternative to chemotherapy for some people with cancer [6] – is consistently found in propolis regardless of geographical location, and thus a good example of an Old Friend. Clearly these essential symbionts will be harvested locally and have a local bias depending on geology, climate and the prevailing mix of plants and fungi; to this extent a honey bee colony is inseparable from its landscape. The perils of moving bees out of their natural range and dissociating them from their original source of symbionts can thus start to be understood. This also highlights the huge negatives of sugar feeding, which not only dislocates the colony from the ebbs and flows of seasonal forage availability but also displaces inputs of valuable Old Friend symbionts. It is the equivalent of the health dangers posed to mankind by a sterile and often medicated Western diet and Prof. Rook’s words are worth repeating: “without appropriate microbial inputs the regulation of the immune system is faulty, and the risk of chronic inflammatory disorders increases”.
As with humans and their Old Friends, the nest cavity and its microbiome have evolved hand in hand – indeed they are inseparable – and the fact that potent anti-inflammatory reagents and antioxidants like Galangin are generally present shows that the propolis-lined nest cavity does indeed have some of the functions of an immune system. There are many reasons why honey bees – given a choice – might prefer nest spaces with small entrances: the colony is easier to defend, heat and humidity retention is made more efficient, and unwanted intrusion by non-family bees can be regulated. But ensuring that the outer protective boundary is as complete as possible also ensures that the space inside can better function against pathogens and allow a better regulation of all the beneficial microbes within – the bacteria, fungi, viruses collectively known as the microbiome – rather in the way that the human immune system can “farm” the microbiota while at the same time exclude pathogens (to quote Prof. Rook again). The constriction (again with propolis) of nest entrances in winter may be part of a strategy to stay warm, but equally it is an efficient way of containing and maintaining this extended synthetic immune system across the different seasons; maintaining this continuously protected environment will be as important for the contributory microbes as it is for the hosts.



(Matt Somerville)
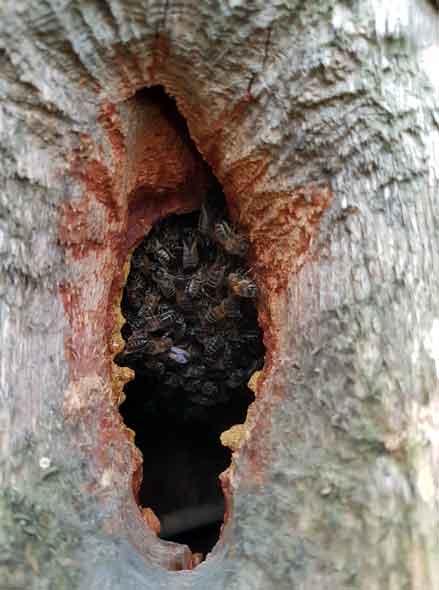
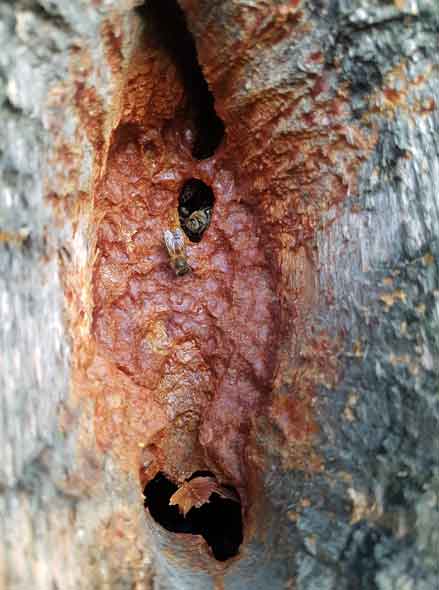
The analogy of the nest envelope with a fully functioning immune system has limitations. The outward-facing element that defines the (super) organism as that which is to be protected is clearly present but the inward-facing portion that looks for potential dangers from within is missing. For this reason, a coping strategy needs to be in place and swarming, leaving aside its prime reproductive function, helpfully provides a regular clear out of pathogens and disease from the nest site. A more extreme reaction, in situations where the colony determines that the current nest site is unsustainable, would be nest abandonment and it is interesting to note the apparent increased frequency of such events being reported. An unoccupied nest site will often be taken over by wax moths and other species that recycle large amounts of the old combs and, with it, disrupt many of the pathogens contained; the propolis wall is left intact and, assuming another species doesn’t move in, the cavity is once again ready for honey bee occupation. Wax moths make a terrible mess and cause economic damage, so modern hive management – obsessed with cleanliness – dictates that they are removed. However, this interference threatens one of the very positive cycles that promote long term colony health and, therefore, survival. Breaking into this cycle, mankind once again perilously upsets the evolved systems that have served bees so well over 16 million years.

Over a decade ago the “Old Friends” hypothesis was expanded to include symbiotic bacteria and parasites as these assist in the function of the immune system: symbiosis means “living together” and is fast replacing many concepts of biological individuality. New technologies, particularly in the area of RNA analysis, have allowed us a much closer look at the microbial world and have revealed layers of complexity and interrelationships that completely challenge the standard view of “individuals” with set boundaries. Our bodies come to be understood as the host for an integrated community of many species living in and around it – a holobiont or “living whole”. There is no doubting that symbiotic microorganisms are fundamental to the biology and ecology of the host by providing and processing vitamins, energy, inorganic or organic nutrients, and participating in defence mechanisms, or indeed by driving the evolution of the host (and the symbiont). The evolutionary aspect is perhaps the most intriguing; thinking of evolutionary pressure as elimination of the weakest (as opposed to survival of the fittest), it may be that the inability of a particular species to survive change is not always driven by a failing in the main host body, but in one of the symbionts. For example, it has been shown that symbionts can buffer honey bee larvae against nutritional stress, meaning that they cope better with the vagaries of forage flows [7]. In other words, the holobiont – the integrated community of species that functions as an apparent whole – becomes the unit of natural selection. This is a strong argument against unnecessary chemical intervention against such parasites as varroa in honey bees; just as with antibiotic use, the course of natural selection is redirected if the challengers to the health of the holobiont are artificially eliminated.
As the immune system ceases to be understood as a stand alone entity, the entire biological individual can be seen to be founded and reliant on the substantial presence of microbiota, to the extent that “individuality” starts to look like a very fuzzy concept indeed. And as we gradually come to realise that our notions of self and individuality are not quite as simple as we had imagined, the honey bee can help teach us that interdependence is actually what defines our place in the world. Each bee – worker, drone or queen – is a holobiont with a large interdependency on a complex ecosystem of symbionts, particularly in the gut which, as with humans, relies on a diverse community of bacteria and anaerobic fungi. These symbionts are part of its anatomy, they enable physiological function and behaviour, and ultimately determine its evolution since, as noted above, selection acts on the whole and the weakest element may just as likely be one of the symbionts as the host. Incredibly, up to 90% of the cells that comprise what we see as a “bee” are in fact bacteria. This defies a simple definition of individual identity at the biological, anatomical or immune levels and, when adding to this the complicating layer of the superorganism (that is the colony), it is difficult not to wonder what actually constitutes an individual honey bee. How can a worker be considered an individual when the reproductive unit of the species is the colony? It cannot even process its primary sources of protein and energy, pollen and nectar, without gut microbes internally and beneficial yeasts and bacteria in the nest it inhabits.
In performing many of the functions of waste processing, nutrient storage and acting as a “womb” at the larval and pupal stages of the bees life, the nest site – not just the superorganism of bees in it – is clearly a form of holobiont itself and since, to a large degree, the symbionts condition and prompt its capabilities, the nest site is – inescapably – also subject to natural selection. Thus the wax combs in the nest are not simply a support or a shelter, but are, like human bones, a living structural component of a fully functioning holobiont, built according to the needs of the colony, with an architecture influenced by the genetics of the inhabitants, the location (in terms of climate and local vegetation) and, of course, the shape and position of the cavity available. Indeed a recent study found that superorganisms even have their own “personalities” – unique temperaments that distinguish them from other colonies in how they respond to threats and accomplish tasks [8]. In this way the design of each nest site varies just as individuals vary; some are doomed to fail, some will survive (at least to the next challenge). The regional influence is mammoth; a colony closely interlinked with its landscape – the microbes in its gut, the scavengers inside the hive, the fungal strains in its water supply, the ebbs and flows of local forage – will clearly have a better chance of survival than one that is not. Even where the local ecotype has been destroyed, bees require the space and time to rebuild their delicate links to the landscape; transfer a colony even a short distance outside its locality and these links are lost. The detriment to health might not be fatal but, in a changing and increasingly sterile environment, still significant enough to cause stress and to interrupt coping mechanisms and any further pressures – pesticide exposure or new parasites such as varroa – will become much more difficult to handle; the straw that breaks the camel’s back. To quote Isobel Grindrod and Stephen Martin, “as [disease] resistance is a population level trait rather than a single colony trait, a resistant colony becomes vulnerable if moved out of its population and could collapse” [2]. As local adaptations are broken and lost, the stress of readapting puts an unwanted and unnecessary burden on the entire population, leading inevitably to such things as increased queen issues and colony collapse being regularly observed.
Since natural selection is the result of differential survival and reproduction rates between colonies, then health and genetics are the keys to maintaining population fitness. The honey bee clearly best suited to a particular environment is the ecotype that has evolved there: the local bee. Self evidently, since it has out-performed all the others, this will be the best survivor in that particular landscape. Here we have the strongest possible argument for keeping honey bees local; by moving them we are not only denying them future access to the genetic pool best suited to their locality, but we are dislocating them from the established sources of the symbionts that make up such a substantial part of their being. We are depriving them of the full ability to benefit from their past evolution; we are messing up the delicate Old Friend relationships that make a honey bee what it is.
REFERENCES
[1] For those wanting to delve further, Prof. Rook has a website here: https://www.grahamrook.net
[2] Grindrod I, Martin SJ. 2021 “Parallel Evolution of Varroa Resistance in Honey Bees: a common mechanism across continents?” (Proc. R. Soc. B 288: 20211375).
[3] A good example of the delicacy of the honey bee gut can be read here: “Nano- and Micro-polystyrene Plastics Disturb Gut Microbiota and Intestinal Immune System in Honeybees” by Wang K, Zhu L, Rao L, Zhao L, Wang Y, Wu X, Zheng H, Liao X. . Sci Total Environ. 2022 Jun 20.
[4] Trans-generational immune priming (TgIP) mechanisms in honey bees is a fascinating subject – see “Trans-generational Immune Priming in Honeybees” Javier Hernández López, Wolfgang Schuehly, Karl Crailsheim, Ulrike Riessberger-Gallé (Proc Biol Sci. 2014 Apr 30;281(1785).
[5] It is worth noting here that honey bees possess only one-third of genes involved in immunity in their genome as compared with mosquitos or small fruit flies, for example. Compensation for this lack of genes has been attributed to the existence of social immunity – see “Social Immunity” by Sylvia Cremer, Sophie A.O. Armitage and Paul Schmid- Hempel (Current Biology 17 R693–R702 August 21 2007).
[6] Heo MY, Sohn SJ, Au WW. “Anti-genotoxicity of Galangin as a Cancer Chemopreventive Agent Candidate” Mutat Res. 2001 May; 488(2).
[7] Parish, A.J., Rice, D.W., Tanquary, V.M. et al. “Honey Bee Symbiont Buffers Larvae Against Nutritional Stress and Supplements Lysine”. SME Journal (June 2022).
[8] “Colony Personality and Plant Health in the Azteca-Cecropia Mutualism” by Peter R Marting, William T Wcislo, Stephen C Pratt (Behavioural Ecology, Volume 29, Issue 1, January/February 2018).